 In the United States alone, the prevalence of obesity is often referred to as an epidemic. Many Americans are overweight, leading to feeling unhappy and unhealthy. Especially true since the turn of the century, healthcare costs increase while overall health decreases; people continue to eat poorly, gain weight, and depend on medications and operations to maintain their health. Statistics can be hard to understand, confusing even those who study nutrition. But the most optimistic numbers state that more than 1/3 of American adults are overweight or obese. Below is a summary of the data presented by the Center for Disease Control (CDC). Here are some shocking childghood obesity facts.
In the United States alone, the prevalence of obesity is often referred to as an epidemic. Many Americans are overweight, leading to feeling unhappy and unhealthy. Especially true since the turn of the century, healthcare costs increase while overall health decreases; people continue to eat poorly, gain weight, and depend on medications and operations to maintain their health. Statistics can be hard to understand, confusing even those who study nutrition. But the most optimistic numbers state that more than 1/3 of American adults are overweight or obese. Below is a summary of the data presented by the Center for Disease Control (CDC). Here are some shocking childghood obesity facts.
- The prevalence of obesity is currently at 42.4% (2018)
- From 2005–2019, the prevalence of obesity increased from 30.5% to 42.4%, and the prevalence of severe obesity increased from 4.7% to 9.2%
- Obesity-related conditions include heart disease, stroke, type 2 diabetes, and certain types of cancer considered to be the leading causes of preventable, premature death
- The estimated annual medical cost of obesity in the United States was $147 billion in 2018 US dollars; the medical cost for people who have obesity was $1,429/year higher than those of normal weight
We can easily make a case for the need to improve health. Diets fail us, fast foods pull us into their offerings without any fear of consequence. Our lack of PA simply adds to the problem. It seems like a good time to view the body and its needs from a different perspective.
Techniques and coaching strategies to guide clients require that we share information. To begin, let’s look at the human form. Once we lay the groundwork for how we are built, we can perhaps better understand how to best care for our bodies. We aim to take a meta-view of the body by breaking down its parts. See the Bonus section for detailed infographics of the body.
The Skeleton
The skeletal system includes not only bones but also cartilage, ligaments, tendons, and other tissues that are essential for everyday functioning. Essentially, anyone who remembers Halloween as a child has had at least some exposure to the human skeleton, and if you’ve been in enough doctor’s offices, you’ve probably seen a high-fidelity or 3-D representation of a skeleton.
The skeleton’s more obvious roles include structural support, locomotion, and protecting the internal organs. Bones produce various types of blood cells and store minerals, particularly calcium, that can be liberated and used elsewhere in the body.
Components of the Skeletal System
The skeletal system includes four basic components, although some sources include a fifth, the joints. The four are the bones, which make up most of the mass of the skeletal system; cartilage, which acts mainly as padding; ligaments, which usually connect bones to bones; and tendons, which join muscles to bones. The system has two divisions, the axial skeleton, and the appendicular skeleton.
Bones are made of a composite tissue, with a high fraction of the mineral calcium. Adult humans have 206 bones; young children may have more because some bones fuse during early maturation. The bones notably provide structural support, giving the body form and rigidity and allowing for walking, running, and more complex movements. But bones are also metabolically active, as the marrow in the interior of bones is where blood cells are made. Bones also feature epithelium, adipose tissue, and nervous tissue.
Tendons connect muscle to bone. They consist of tightly packed bundles of the protein collagen arranged in parallel. They are usually encased in fluid-filled sheaths when they abut other tissues, to decrease friction and increase padding. They act to focus a muscle’s effort on a physically small portion of a bone, better-allowing muscles to serve as levers.
Ligaments connect bones. They do so in a relaxed manner, without any pulling between them since no muscles are involved. They are similar in composition to tendons, although the fibers they are assembled from are often more variable. They sometimes have a degree of elasticity but are not considered “stretchy.”
Cartilage is found in joints between bones and is made of material intermediate in firmness between bones at the hard end and tendons and ligaments at the softer end. It contains both collagen and chondroitin sulfate. Its appearance is sponge-like, with pores called lacunae between the solid portions. It comes in many forms, the most common being hyaline cartilage, elastic cartilage, fibrocartilage, and calcified cartilage.
Components of the Axial Skeleton
The axial skeleton is so named because it includes the 80 bones along the long axis of the body above the pelvis. From top to bottom, it includes the skull, mandible (lower jaw), hyoid bone (under the chin), vertebral column, ribs, and sternum.
The skull alone contains 22 bones. Seven additional bones, including the ossicles of the inner ear and the hyoid, are associated with the skull. The skull serves to house and protects the brain. It is connected to the top of the vertebral column.
The vertebral column includes 24 bones, along with the sacrum at the base and the coccyx (tail bone) at the lower end of the sacrum. The vertebrae protect the spinal cord and allow for upright posture.
Humans have 12 pairs of ribs, which serve to guard the heart, lungs, and other internal organs of the thorax. Between them in the front of the body is the sternum or breastbone.
Components of the Appendicular Skeleton
The appendicular skeleton, so named because it includes the body’s appendages (arms and legs in humans), features 126 bones.
The long bones of the arm include the humerus of the upper arm and the radius and ulna of the lower arm. The long bones of the leg are the femur, or thigh bone, and the tibia (shin bone) and fibula of the lower leg. The primary pelvic bone is the ilium; the points you can feel at the top of each of your hips are called the iliac crests.
The hands (26 bones each) and the feet (27 apiece) together include over half of the bones in the human body – 106 out of the total 206 listed in most textbooks. The hands and feet each have 14 phalanges, which are the small joints of the fingers and toes (each finger has three, the thumb two; the second through fifth toes also have three each, while the big toe, like the thumb, includes two). Each hand and each foot has five bones that connect phalanges to the wrist and ankle bones respectively; these bones form the scaffolding of the palms (the metacarpals) and the arches of the feet (the metatarsals). Finally, each wrist has eight bones, while each ankle includes seven.
Skeletal System Organs and Tissues
Each bone is actually an organ in its own right, and each of our long bones includes a diaphysis (shaft) and an epiphysis at each end.
In addition to the long bones of the appendicular skeleton, the body includes various other types of bones. One of these is short bones, which include the irregular bones of the wrist and ankle. Flat bones, which include the sternum, the ilium, and the skull bones, are mostly protective in function, and in the pelvis, their considerable surface area allows for many different muscles to attach to the same general part of the skeleton. The vertebrae and the hyoid are irregular bones, which have functions determined by their location in the body. Finally, there are the sesamoid bones, which mainly protect tendons and most notably include the patellae (kneecaps).
The marrow in the middle of bones comes in two forms, yellow and red. Yellow bone marrow contains adipose (fat) tissue, which can be released to serve as an energy source for other tissues. Red bone marrow is where blood cells are manufactured, a process called hematopoiesis. Red blood cells, white blood cells, and platelets are all produced in this type of marrow.
Skeletal System Function
Without the joints, the skeletal system would not be able to engage in the numerous refined functions of movement that give vertebrate animals their form and function. Joints come in three types:
Synarthroses are immovable joints and include the sutures between the skull bones, the points of contact between the teeth and the mandible, and the joint that is located between the topmost pair of ribs and the sternum. Amphiarthroses are joints that permit a small amount of movement. These include the joint between the tibia and the fibula just above the foot and the pubic symphysis that joins the two sides of the pelvis in the front of the body at the waist. Diarthroses are joints that allow full movement and include many of the joints in the upper and lower limbs, such as the elbow, shoulder, and ankle joints. Learn more about joint function and dysfunction.
The minerals in bones, including calcium and phosphorus, can be released from bone into the bloodstream for the maintenance of metabolic functions as calcium ions participate in muscle contraction, and phosphates are a critical component of DNA and molecules important in energy transfer and release.
- Structure
Like the steel framework of a building, the functions of the skeleton and the bones are to provide rigidity, which gives the body shape and supports the weight of the muscles and organs. Without this structure, the body would collapse in on itself, compressing the lungs, heart, and other organs–impairing their function. The rigid structure of the skeletal system also allows it to perform another one of the 5 functions of the skeletal system: movement.
- Movement
There are three major systems involved in the mechanics of movement:
- Nervous system
- Muscular system
- Skeletal system
The nervous system sends the electrical impulses that activate the muscles, the skeletal system provides the levers and anchors for the muscles to pull against. All skeletal muscles have an origin and insertion point.
The origin is the proximal anchor and is closest to the center or axiom of the body’s skeletal frame. This is the portion of bone that remains mostly immobile while the muscle works. The insertion is the bone that moves as the muscle works, which is one of the main functions of the skeleton. So, for example, in the case of the biceps, the upper arm and shoulder are the origins (anchor) and the bones of the forearm are the insertion.
Interestingly, the amount of power the muscle needs is directly related to the length of the bone (or lever) and where it is attached.
- Protection
Arguably the most important of the 5 functions of the skeletal system is protection. The most obvious example of the functions of the skeleton’s protective properties is the human skull. The vertebrae and ribs also have protective functions by encasing delicate structures like the spinal cord, heart, and lungs. The rib cage not only surrounds the organs of respiration, but it’s also very flexible and is constructed to expand and contract with each breath.
Human vertebrae are all irregularly shaped bones in the human body that provide both protection and flexibility for movement. There are also fibrous disks between each vertebra, which provide shock absorption.
- Blood Cell Production
Red and white blood cells are made in the red marrow of bones. At birth and in early childhood, all bone marrow is red. As we age, about half of the body’s marrow turns to yellow marrow – which is composed of fat cells. In an adult human, most of the long bones contain yellow marrow, and the red marrow is only found in the flat bones of the hip, skull and shoulder blades, the vertebrae, and at the ends of the long bones.
- Storage
The body uses calcium and phosphorous for bodily processes like muscle contraction. Some of those minerals are found in our diet, but they are also taken from the bones in the human body. When the body needs calcium, if there isn’t a ready supply in the blood, the endocrine system releases hormones that initiate the process of taking calcium from bone and releasing it into the bloodstream. When there is a surplus of blood calcium, it’s put back into the bones.
This is precisely why dietary calcium and vitamin D are important. The body uses calcium constantly and, if there isn’t enough calcium in the diet, it will consistently take calcium from the bone to compensate – leading to osteoporosis. Having enough dietary calcium ensures that there is enough calcium for bodily functions and replenishes the backup stores in the bone.
The Muscular System
Muscles of the human body work to engage and control the skeletal system that is under voluntary control. This includes those use for movement, posture, and balance. The human muscle system is often taught by dividing it into striated muscle (or skeletal muscle), smooth muscle, and cardiac muscle.
A Meta-view of Human Musculature
Smooth muscle is under involuntary control and is found in the walls of blood vessels and structures such as the urinary bladder, the intestines, and the stomach. Cardiac muscle makes up the mass of the heart and is responsible for the rhythmic contractions of that vital pumping organ; it is also under involuntary control. With very few exceptions, the arrangement of smooth muscle and cardiac muscle in humans is identical to the arrangement found in other vertebrate animals.
Smooth muscle and cardiac muscle and the physiology of muscle contraction are described later.
Muscle Groups and Their Actions
The following sections provide a basic framework for the understanding of human muscular anatomy, with descriptions of the large muscle groups and their actions. It is important to know that muscle groups work in a coordinated fashion (synergistically) to control the movements of the human body.
The Neck
The motion of the neck is described in terms of rotation, flexion, extension, and side bending (i.e., the motion used to touch the ear to the shoulder). The direction of the action can be ipsilateral, which refers to movement in the direction of the contracting muscle, or contralateral, which refers to movement away from the side of the contracting muscle. Here’ show to work with neck injuries in yoga.
Rotation is one of the most important actions of the cervical (neck) spine.
Rotation is accomplished primarily by the sternocleidomastoid muscle, which bends the neck to the ipsilateral side and rotates the neck contralaterally. Together, the sternocleidomastoid muscles on both sides of the neck act to flex the neck and raise the sternum to assist in forced inhalation. The anterior and middle scalene muscles, which also are located at the sides of the neck, act ipsilaterally to rotate the neck, as well as to elevate the first rib. The splenius capitis and splenius cervicis, which are in the back of the neck, work to rotate the head.
Lateral flexion is also an important action of the cervical spine. The sternocleidomastoid muscles are involved in cervical side bending. The posterior scalene muscles, located on the lower sides of the neck, ipsilaterally bend the neck to the side and elevate the second rib. The splenius capitis and splenius cervicis also assist in neck side bending. The erector spinae muscles (iliocostalis, longissimus, and spinalis) are large, deep muscles that extend the length of the back. All three act to bend the neck in multiple planes.
Neck flexion refers to the motion used to touch the chin to the chest. It is accomplished primarily by the sternocleidomastoid muscles, with assistance from the longus colli and the longus capitis, which are found in the front of the neck. Neck extension is the opposite of flexion and is accomplished by many of the same muscles that are used for other neck movements, including the splenius cervicis, splenius capitis, iliocostalis, longissimus, and spinalis muscles.
The Back
The back contains the origins of many of the muscles that are involved in the movement of the neck and shoulders. Also, the axial skeleton that runs vertically through the back protects the spinal cord, which innervates almost all the muscles in the body.
Multiple muscles in the back function specifically in movements of the back. The erector spinae muscles, for example, extend the back (bend it backward) and side bend the back. The semispinalis dorsi and semispinalis capitis muscles also extend the back. The small muscles of the vertebrae (the multifidi and rotators) help rotate, extend, and bend the back laterally. The quadratus lumborum muscle in the lower back bends the lumbar spine laterally and aids in the inspiration of air through its stabilizing effects at its insertion at the 12th rib (the last of the floating ribs). The scapula (shoulder blade) is elevated by the trapezius muscle, which runs from the back of the neck to the middle of the back, by the rhomboid major and rhomboid minor muscles in the upper back, and by the levator scapulae muscle, which runs along the side and back of the neck.
The Shoulder
The shoulder is a complex ball-and-socket joint comprising the head of the humerus, the clavicle (collarbone), and the scapula. The shoulder’s main motions are flexion, extension, abduction, adduction, internal rotation, and external rotation.
Shoulder flexion is a movement of the shoulder in a forward motion. An example of shoulder flexion can be seen when reaching forward to grasp an object. That action is accomplished primarily by the combined actions of the deltoid muscle in the uppermost extent of the arm, the pectoralis major muscle in the chest, the coracobrachialis muscle on the inside of the upper arm, and the biceps brachii muscles on the front of the upper arm.
Extension of the shoulder is opposite to flexion. Pure shoulder extension is the movement of the arm directly behind the body, as in receiving a baton in a relay race. That movement is accomplished by the actions of the deltoid muscle, the latissimus dorsi muscle in the back, the teres major muscle in the armpit area, and the triceps muscle in the back of the upper arm. The triceps, as the name suggests, consists of three heads that originate from different surfaces but share the same insertion at the olecranon process of the ulna (a bone in the forearm); the three heads together act to extend the elbow.
Shoulder adduction and abduction serve to lower the arm toward and lift the arm away from the body, respectively. They can be visualized by picturing someone doing jumping jacks. Adduction is accomplished primarily by the pectoralis major, latissimus dorsi, teres major, triceps, and coracobrachialis. The deltoid and the supraspinatus, a muscle that runs along the scapula in the back, are the two main abductors of the shoulder.
An example of an external rotation of the shoulder is seen in a tennis backhand stroke. External rotation is attributed primarily to the deltoid, the teres minor in the armpit area, and the infraspinatus muscle, which covers the scapula. Internal rotation of the shoulder is the opposite of external rotation. An example is the shoulder movement that occurs when reaching into a back pocket. That movement is achieved through the coordinated action of the pectoralis major, latissimus dorsi, deltoid, teres major, and subscapularis muscles. (The subscapularis is a deep muscle situated on the anterior, or front-facing, surface of the scapula.)
The teres minor, subscapularis, supraspinatus, and infraspinatus muscles together form the rotator cuff, which stabilizes the humeral head (the ball portion of the ball-and-socket shoulder joint). The muscles of the rotator cuff are common sites of injury in adults, particularly among people who perform overhead motions repeatedly (e.g., throwing a baseball or painting a ceiling). Several of the rotator cuff muscles have tendons that run under the acromion, a bony prominence at the distal end of the scapula. (The term distal describes a relative position away from the center of the body; it often is contrasted with the term proximal, which describes a relative position near to the center of the body.) The position of the tendons and the subacromial bursae (fluid-filled sacs located beneath the acromion) leaves them vulnerable to compression and pinching, which can result in an injury known as shoulder impingement syndrome.
The Arm
In addition to aiding the movement of the shoulder, the muscles of the upper arm produce various movements of the forearm. For example, the primary muscles involved in forearm flexion, in which the angle formed at the elbow becomes smaller (i.e., the hand moves closer to the shoulder), are the biceps brachii, the brachialis (situated beneath the biceps brachii in the upper arm), and the brachioradialis (the origin of which is on the humerus). Minor contributions to forearm flexion are provided by the coracobrachialis and by flexor muscles situated in the anterior compartment of the forearm.
Extension of the forearm increases the angle at the elbow, moving the hand away from the shoulder. That action is accomplished primarily by the triceps brachii. Other muscles that make minor contributions to forearm extension include the extensor muscles of the posterior compartment of the forearm (the side of the forearm that is contiguous with the back of the hand; also known as the extensor compartment), including the extensor carpi radialis longus, the extensor carpi radialis brevis, the extensor digitorum, the extensor carpi ulnaris, and the anconeus.
The Wrist
Wrist flexion refers to the movement of the wrist that draws the palm downward. That action is carried out by the flexor carpi radialis, the flexor carpi ulnaris, the flexor digitorum superficialis, the flexor digitorum profundus, and the flexor pollicis longus. Here are common dysfunctions of the wrist.
Wrist extension, by contrast, shortens the angle at the back of the wrist. The muscles responsible for that action are the extensor carpi radialis longus and the extensor carpi radialis brevis, which also abduct the hand at the wrist (move the hand in the direction of the thumb, or first digit); the extensor digitorum, which also extends the index to the little finger (the second to fifth digits); the extensor digiti minimi, which also extends the little finger and adducts the hand (moves the hand in the direction of the little finger); and the extensor carpi ulnaris, which also adducts the hand. Other small muscles that cross the wrist joint may add to wrist extension, but they do so to only a small degree.
Supination is the rotation of the wrist that brings the palm facing up. The supinator muscle in the posterior compartment acts to supinate the forearm. The biceps brachii also adds to supination. Pronation is the opposing action, in which the wrist is rotated so that the palm is facing down. The pronator quadratus, a deep muscle in the anterior compartment, along with the pronator teres, pronates the forearm.
The Hand
The hand is a complex structure that is involved in fine motor coordination and complex task performance. Its muscles generally are small and extensively innervated. Even simple actions, such as typing on a keyboard, require a multitude of precise movements to be carried out by the hand muscles. Because of that complexity, the following paragraphs cover only the primary action of each hand muscle.
Several muscles that originate at the posterior surface of the ulna or the radius (the other bone in the forearm) have their actions in the hand. Those include the abductor pollicis longus, which abducts and extends the thumb; the extensor pollicis brevis, which extends the metacarpophalangeal (MCP) joint of the thumb; the extensor pollicis, which extends the distal phalanx (finger bone) of the thumb; and the extensor indicis, which extends the index finger at the MCP joint. (MCP joints are located between the metacarpal bones, which are situated in the hand, and the phalanges, which are the small bones of the fingers.)
Although several of the muscles that move the hand have their origins in the forearm, there are many small muscles of the hand that have both their origin and their insertion within the hand. Those are referred to as the intrinsic muscles of the hand. They include the palmaris brevis, which assists with grip; the umbricals, which flex the MCP joints and extend the interphalangeal joints (IPs; the joints between the phalanges) of the fingers; the palmar interossei, which adduct the fingers toward the middle finger (the third digit); and the dorsal interossei, which abduct the fingers away from the middle finger. All the interossei flex the MCP joints and extend the IP joints.
The thenar eminence is located on the palm side of the base of the thumb and is composed of three muscles, the abductor pollicis brevis, the flexor pollicis brevis, and the opponens pollicis, all of which are innervated by the median nerve. The abductor pollicis brevis abducts the thumb; the flexor pollicis brevis flexes the MCP joint of the thumb; and the opponens pollicis acts to oppose the thumb to the other fingers. The adductor pollicis, which is not part of the thenar eminence, acts to adduct the thumb.
The hypothenar enimence is located on the palm side of the hand below the little finger. It contains three muscles that are innervated by the deep branch of the ulnar nerve. The abductor digiti minimi abducts the little finger. The flexor digiti minimi flexes the little finger. The opponens digiti minimi opposes the little finger with the thumb.
The Abdomen
There are three muscular layers of the abdominal wall, with a fourth layer in the middle anterior region. The fourth layer in the mid-region is the rectus abdominis, which has vertically running muscle fibers that flex the trunk and stabilize the pelvis. To either side of the rectus abdominis are the other three layers of abdominal muscles. The deepest of those layers is the transversus abdominis, which has fibers that run perpendicular to the rectus abdominus; the transversus abdominis acts to compress and support the abdomen and provides static core stabilization. The internal oblique layers run upward and forward from the sides of the abdomen, and the external oblique layers, which form the outermost muscle layers of the abdomen, run downward and forward. The internal oblique layers act in conjunction with the external oblique on the opposite side of the body to flex and rotate the trunk toward the side of the contracting internal oblique.
The Hip
The hip joint is a complex weight-bearing ball-and-socket joint that can sustain considerable load. The socket of the joint is relatively deep, allowing for stability but sacrificing some degree in range of motion. The movements described in this section include flexion, extension, abduction, and adduction.
Hip flexion is the hip motion that brings the knee toward the chest. The major muscles of hip flexion include the iliopsoas, which is made up of the psoas major, psoas minor, and iliacus. Together, those muscles act mainly to flex the hip, but they also contribute to abdominal flexion and hip stabilization. Other hip flexors include the sartorius, the rectus femoris, the pectineus, and the gracilis. The sartorius also contributes to external hip rotation and knee extension and abduction, and the rectus femoris also acts in knee extension. The pectineus is also involved in hip adduction and internal rotation.
Hip extension is accomplished primarily by the muscles of the posterior thigh and buttocks, which when contracted serve to move the thigh from a flexed position toward the midline of the body or the trunk of the body from a bent position toward a more-erect posture. Hip extension is accomplished mostly by the gluteus maximus, the biceps femoris (which is divided into two heads, the long head, and the short head), the semitendinosus, and the semimembranosus. A minor contribution is also provided by the adductor magnus and other small pelvic muscles.
The movement of adduction is used to describe a direction of limb motion that serves to take the limb from a lateral position to its more-axial alignment. During a jumping-jack exercise, for example, the abduction of the leg occurs when it is moved away from the midline and adduction when it is moved back toward the midline. The main abductors of the hip are the gluteus medius, gluteus minimus, and tensor fascia lata. These three muscles also serve to internally rotate the thigh in an extended position and externally rotate the thigh in the flexed position. Another minor contributor is the piriformis. The main hip adductors are the adductor magnus, the adductor brevis, and the adductor longus. A minor contribution to hip adduction is performed by the pectineus and the gracilis.
The Upper Leg and Knee
Extension of the knee is accomplished by a group of muscles collectively referred to as the quadriceps femoris, which increases the angle of the knee, bringing the lower leg into a straight position. Knee extension is used in the forward, swing phase of the gait and is integral in movements such as kicking. The quadriceps femoris group includes the vastus medius, vastus lateralis, vastus intermedius, and rectus femoris. A minor contribution to knee extension is provided by the sartorius.
Knee flexion refers to the bending of the knee from the straight position. The muscles that perform that action oppose those of knee extension and are referred to as the hamstring muscles. The hamstring muscles are situated in the back of the thigh and include the biceps femoris, the semitendinosus, and the semimembranosus. Small contributions to knee flexion are made by the gastrocnemius muscle in the back of the calf and by several small muscles that cross the knee joint posteriorly. This answers many questions about exercises involving the knees.
The Lower Leg and Foot
The muscles of the lower leg and foot are complex and work in many planes. Their actions depend on whether the person is bearing weight, as well as on the position of the foot. The following paragraphs provide a brief overview of the actions of the muscles of the lower leg and foot.
Dorsiflexion refers to ankle flexion in the direction of the dorsum, or anterior surface of the foot (the surface of the foot viewed from above). Dorsiflexion is accomplished by several muscles, including the tibialis anterior, which in addition to dorsiflexion also inverts the foot (tilts the foot toward the midline), stabilizes the foot when striking the ground, and locks the ankle when kicking. The extensor digitorum longus (EDL) also acts in dorsiflexion and functions to extend the last four toes. In addition to the EDL, some individuals also have a muscle called the peroneus tertius (fibularis tertius), which participates to a limited extent in dorsiflexion and eversion of the foot (tilting of the foot away from the midline). The extensor hallucis longus primarily acts in big toe (hallux) dorsiflexion, but it also acts to dorsiflex, as well as weakly invert, the ankle. Here’s more information about what trainers and coaches sould know about common lower leg and foot injuries.
Plantar flexion refers to the flexion of the ankle in the direction of the sole. Most of the ankle plantarflexion is performed by the large calf musculature, including the gastrocnemius and the soleus, which lies just behind the gastrocnemius. It is generally accepted that those are two distinct muscles; however, there is some debate as to whether the gastrocnemius and the soleus are two parts of the same muscle.
Other muscles of the lower leg and foot include The Plantaris, which runs obliquely between the gastrocnemius and the soleus; the flexor hallucis longus, which contributes to ankle flexion but is involved primarily in big toe flexion; the flexor digitorum longus, which also flexes the second to fifth toes; the peroneus longus, which flexes the ankle and everts the foot; and the peroneus brevis, which is involved in plantarflexion and eversion of the foot.
Intrinsic muscles of the foot arise in the foot and do not cross the ankle joint. Hence, their action is confined to the foot. The intrinsic muscles of the foot include the abductor hallucis, which abducts the big toe; the flexor digitorum brevis, which flexes the second to fifth toes; the abductor digiti minimi, which abducts and flexes the fifth toe; the quadratus plantae, which assists in toe flexion; the lumbricals, which flex the metatarsophalangeal (MTP) joints and extend the distal IP and proximal IP joints of the toes; the flexor hallucis brevis, which flexes the big toe; and the adductor hallucis, which flexes and contracts the big toe. The adductor hallucis has two heads, the oblique head and the transverse head, which share an insertion on the lateral (outer) side of the base of the proximal phalanx of the big toe. The oblique head arises from the base of the second to fourth metatarsal bones, and the transverse head arises from the ligaments of the MTP joints of the third to fifth toes. The flexor digiti minimi brevis extends and adducts the fifth toe. The dorsal interossei abduct the toes, and The Plantar interossei adduct the toes.
The arrangement of striated muscle in humans conforms to the basic plan seen in all pronograde quadrupedal vertebrates and mammals (that is, all vertebrates and mammals that assume a horizontal and four-legged posture). The primates (the order of mammals to which humans belong) inherited the primitive quadrupedal stance and locomotion, but since their appearance in the Late Cretaceous Period some 65 million years ago, several groups have modified their locomotor system to concentrate on the use of the arms for propulsion through the trees. The most extreme expression of that skeletal adaptation in living primates is seen in the modern gibbon family. Their forelimbs are relatively elongated; they hold their trunk erect; and, for the short periods that they spend on the ground, they walk only on their hind limbs (in a bipedal fashion).
The Cardiovascular System
Every day, thousands of Americans die from cardiovascular disease. On a global level, Cardiovascular diseases (CVD’s) are the number 1 cause of death globally, taking an estimated 17.9 million lives each year. Four out of 5 CVD deaths are due to heart attacks and strokes, and one-third of these deaths occur prematurely in people under 70 years of age. Why are we so unhealthy and overweight? Why do we see increasing numbers from chronic health concerns, such as heart disease, obesity, diabetes, reproductive issues, and depression? Why do we need so much medical attention and medication? In a word, it is ‘lifestyle’.
Which exercises are best to improve heart health and strengthen the cardiovascular system?
Preventing Heart Disease
When experts talk about preventing heart disease, they usually refer to one of three types: secondary, primary, and primordial prevention. All three have similar elements, but different starting times and different effects. Some, the CHNC can address; others are more for those with medical degrees.
Primary prevention. Primary prevention aims to keep an individual at risk of heart disease from having a first heart attack or stroke, needing angioplasty or surgery, or developing some other form of heart disease. Primary prevention is usually aimed at people who already have developed cardiovascular risk factors, such as high blood pressure or high cholesterol. As with secondary prevention, primary prevention focuses on controlling these risk factors by making healthy lifestyle changes and, if needed, taking medications. That said, the appearance of worrisome cardiovascular risk factors means that inflammation, atherosclerosis, and/or endothelial dysfunction are already at work and, in most cases, aren’t reversible.
Secondary prevention. These efforts are started after someone has a heart attack or stroke, undergoes angioplasty or bypass surgery, or develops some other form of heart disease. It involves taking medications like aspirin and/or a cholesterol-lowering statin, quitting smoking and losing weight if needed, exercising more, and following a healthy diet. Although secondary prevention may sound like “closing the barn door after the horse has gone,” it isn’t. These steps can prevent a second heart attack or stroke, halt the progression of heart disease, and prevent early death. It may be obvious, but the number one killer of individuals who survive a first heart attack is a second heart attack.
Primordial prevention. The word “primordial” means existing from the beginning. Primordial prevention involves working to prevent inflammation, atherosclerosis, and endothelial dysfunction from taking hold, and thus prevent risk factors such as high blood pressure, high cholesterol, excess weight, and ultimately cardiovascular events. Once rarely discussed, primordial prevention is now the cornerstone of the American Heart Association’s definition of ideal heart health and efforts to help people achieve it. As its name implies, the sooner one can start practicing primordial prevention—ideally from childhood on—the more likely they are to achieve it and protect themselves from heart disease.
Steps for the Primordial Prevention of Heart Disease
Four key lifestyle steps can dramatically reduce the chances of developing cardiovascular risk factors and ultimately heart disease:
- Not smoking
- Maintaining a healthy weigh
- Exercising
- Following a healthy diet – healthy eating should not be optional
The last of these steps is where the Certified Holistic Nutrition Coach can have the greatest impact, and many believe that the best diet for preventing heart disease is one that is full of fruits and vegetables, whole grains, nuts, fish, poultry, and vegetable oils; it includes alcohol in moderation, with less red and processed meats. Refined carbohydrates, foods, and beverages with added sugar, sodium, and foods with trans-fat are the obvious items to avoid.
How do we know this? Research on people with diets consistent with these dietary recommendations may expect to see a 31% lower risk of heart disease, a 33% lower risk of diabetes, and a 20% lower risk of stroke, according to the Harvard School of Public Health.
A randomized controlled trial found that a Mediterranean diet supplemented with extra-virgin olive oil or nuts, both rich sources of unsaturated fat, reduced the incidence of major cardiovascular events amongst patients with cardiovascular disease over a 4.8-year follow-up period.
Cardiovascular diseases (CVDs) are a group of disorders of the heart and blood vessels and they include:
- coronary heart disease – disease of the blood vessels supplying the heart muscle
- cerebrovascular disease – disease of the blood vessels supplying the brain
- peripheral arterial disease – disease of blood vessels supplying the arms and legs
- rheumatic heart disease – damage to the heart muscle and heart valves from rheumatic fever, caused by streptococcal bacteria
- congenital heart disease – malformations of heart structure existing at birth
- deep vein thrombosis and pulmonary embolism – blood clots in the leg veins, which can dislodge and move to the heart and lungs
This study highlighted that low-fat diets are not beneficial to heart health and that incorporating healthy fats – such as those included in the Mediterranean diet – can improve heart health and weight loss.
There isn’t one Mediterranean diet, as this eating style considers the different foods, eating patterns, and lifestyles in multiple countries that border the Mediterranean region. However, some similarities define a Mediterranean eating pattern, including high intake of olive oil, nuts, vegetables, fruits, and cereals; moderate intake of fish and poultry; low intake of dairy products, red meat, processed meats, and sweets. Wine is encouraged in moderation, consumed with meals.
More of the body’s systems that require sound nutritional intake will continue to be part of this training. We now turn to look at the lymphatic system.
The Lymphatic Drainage System
The lymphatic system can be thought of as an efficient draining system. As blood circulates through the body, blood plasma transfers into tissues through the thin walls of the capillaries. The portion of blood plasma that escapes is called interstitial or extracellular fluid, containing oxygen, glucose, amino acids, and other nutrients needed by tissue cells. Although most of this fluid seeps immediately back into the bloodstream, a percentage of it, along with the particulate matter, is left behind. The lymphatic system removes this extra fluid and these materials from tissues, returning them via the lymphatic vessels to the bloodstream. This is to prevent a fluid imbalance that would result in the organism’s death.
The fluid and proteins within the tissues begin their journey back to the bloodstream by passing into tiny lymphatic capillaries that infuse almost every tissue of the body. Only a few regions, including the epidermis of the skin, the mucous membranes, the bone marrow, and the central nervous system, are free of lymphatic capillaries, whereas regions such as thelungs, gut, genitourinary system, and dermis of the skin are densely packed with these vessels. Once within the lymphatic system, the extracellular fluid, which is now called lymph, drains into larger vessels called the lymphatics. These vessels converge to form one of two large vessels called lymphatic trunks, which are connected to veins at the base of the neck. One of these trunks, the right lymphatic duct, drains the upper right portion of the body, returning lymph to the bloodstream via the right subclavian vein. The other trunk, the thoracic duct, drains the rest of the body into the left subclavian vein. Lymph is transported along with the system of vessels by muscle contractions, and valves prevent lymph from flowing backward. The lymphatic vessels are punctuated at intervals by small masses of lymph tissue, called lymph nodes, that remove foreign materials such as infectious microorganisms from the lymph filtering through them.
Role in Immunity
In addition to serving as a drainage network, the lymphatic system helps protect the body against infection by producing white blood cells called lymphocytes, which help rid the body of disease-causing microorganisms. The organs and tissues of the lymphatic system are the major sites of production, differentiation, and proliferation of two types of lymphocytes—the T lymphocytes and B lymphocytes, also called T cells and B cells. Although lymphocytes are distributed throughout the body, it is within the lymphatic system that they are most likely to encounter foreign microorganisms. This is how to build a strongimunse system.
Lymphoid Organs
The lymphatic system is commonly divided into the primary lymphoid organs, which are the sites of B and T cell maturation, and the secondary lymphoid organs, in which further differentiation of lymphocytes occurs. Primary lymphoid organs include the thymus and bone marrow. In humans, the thymus and bone marrow are the key players in immune function. All lymphocytes derive from stem cells in the bone marrow. Stem cells destined to become B lymphocytes remain in the bone marrow as they mature, while prospective T cells migrate to the thymus to undergo further growth. Mature B and T lymphocytes exit the primary lymphoid organs and are transported via the bloodstream to the secondary lymphoid organs, where they become activated by contact with foreign materials, such as particulate matter and infectious agents, called antigens in this context.
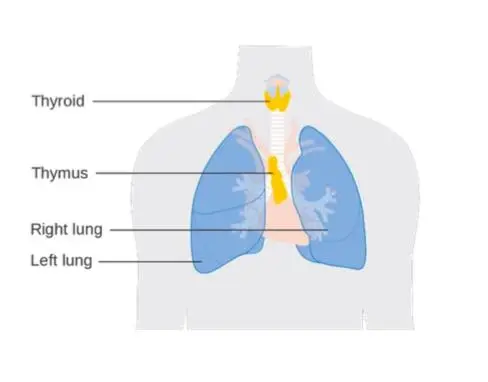
Thymus
The thymus is located just behind the sternum in the upper part of the chest. It is a bilobed organ that consists of an outer, lymphocyte-rich cortex and an inner medulla. The differentiation of T cells occurs in the cortex of the thymus. In humans, the thymus appears early in fetal development and continues to grow until puberty, after which it begins to shrink. The decline of the thymus is thought to be the reason T-cell production decreases with age.
In the cortex of the thymus, developing T cells, called thymocytes, come to distinguish between the body’s components, referred to as “self,” and those substances foreign to the body, called “nonself.” This occurs when the thymocytes undergo a process called positive selection, in which they are exposed to self- molecules that belong to the major histocompatibility complex (MHC). Those cells capable of recognizing the body’s MHC molecules are preserved, while those that cannot bind these molecules are destroyed. The thymocytes then move to the medulla of the thymus, where further differentiation occurs. There, thymocytes that can attack the body’s tissues are destroyed in a process called negative selection.
Positive and negative selection destroy a great number of thymocytes; only about 5 to 10 percent survive to exit the thymus. Those that survive leave the thymus through specialized passages called efferent (outgoing) lymphatics, which drain to the blood and secondary lymphoid organs. The thymus has no afferent (incoming) lymphatics, which supports the idea that the thymus is a T-cell factory rather than a rest stop for circulating lymphocytes.
Bone Marrow
In mammals, the primary organ for B-lymphocyte development is the bone marrow, although the prenatal site of B-cell differentiation is the fetal liver. Unlike the thymus, the bone marrow does not atrophy at puberty, and therefore there is no concomitant decrease in the production of B lymphocytes with age.
Secondary Lymphoid Organs
Secondary lymphoid organs include the lymph nodes, spleen, and small masses of lymph tissue such as Peyer’s patches, the appendix, tonsils, and selected regions of the body’s mucosal surfaces (areas of the body lined with mucous membranes). The secondary lymphoid organs serve two basic functions: they are a site of further lymphocyte maturation, and they efficiently trap antigens for exposure to T and B cells.
Lymph Nodes
The lymph nodes, or lymph glands, are small, encapsulated bean-shaped structures composed of lymphatic tissue. Thousands of lymph nodes are found throughout the body along the lymphatic routes, and they are especially prevalent in areas around the armpits (axillary nodes), groin (inguinal nodes), neck (cervical nodes), and knees (popliteal nodes). The nodes contain lymphocytes, which enter from the bloodstream via specialized vessels called the high endothelial venules. T cells congregate in the inner cortex (paracortex), and B cells are organized in germinal centers in the outer cortex. Lymph, along with antigens, drains into the node through afferent (incoming) lymphatic vessels and percolates through the lymph node, where it encounters and activates lymphocytes. Activated lymphocytes, carried in the lymph, exit the node through the efferent (outgoing) vessels and eventually enter the bloodstream, which distributes them throughout the body.
Spleen
The spleen is found in the abdominal cavity behind the stomach. Although structurally like a lymph node, the spleen filters blood rather than lymph. One of its main functions is to bring blood into contact with lymphocytes. The functional tissue of the spleen is made up of two types of cells: the red pulp, which contains cells called macrophages that remove bacteria, old blood cells, and debris from the circulation process; and surrounding regions of white pulp, which contain great numbers of lymphocytes. The splenic artery enters the red pulp through a web of small blood vessels, and blood-borne microorganisms are trapped in this loose collection of cells until they are gradually washed out through the splenic vein. The white pulp contains both B and T lymphocytes
Mucosa-associated Tissues
Another group of important secondary lymphoid structures is the mucosa- associated lymphoid tissues. These tissues are associated with mucosal surfaces of almost any organ, but especially those of the digestive, genitourinary, and respiratory tracts, which are constantly exposed to a wide variety of potentially harmful microorganisms and therefore require their system of antigen capture and presentation to lymphocytes. For example, Peyer’s patches, which are mucosa- associated lymphoid tissues of the small intestine, sample passing antigens and expose them to underlying B and T cells. Other, less-organized regions of the gut also play a role as secondary lymphoid tissue.
Diseases of the Lymphatic System
A group of secondary lymphoid organs provides a system of redundancy for antigen sampling by the cells of the immune system. Removal of the spleen selected lymph nodes, tonsils, or appendix does not generally result in an excessive increase in disease caused by pathogenic microorganisms. However, the importance of the primary lymphoid organs is clear. For example, two autoimmune diseases, DiGeorge syndrome, and Nezelof disease result in the failure of the thymus to develop and in the subsequent reduction in T-cell numbers, and removal of the bursa from chickens results in a decrease in B-cell counts. The destruction of bone marrow also has devastating effects on the immune system, not only because of its role as the site of B-cell development but also because it is the source of the stem cells that are the precursors for lymphocyte differentiation.
The Lungs
Among air-breathing vertebrates, lungs refer to either of the two largeorgans of respiration located in the chest cavity and responsible for adding oxygen to and removing carbon dioxide from the blood. In humans, each lung is encased in a thin membranous sac called the pleura, and each is connected with the trachea (windpipe) by its main bronchus (large air passageway) and with the heart by the pulmonary arteries. The lungs are soft, light, spongy, elastic organs that normally, after birth and always contain some air. If healthy, they will float in water and crackle when squeezed; diseased lungs sink. Here’s how the lungs work during exercise.
The Mechanics of Breathing
Air moves in and out of the lungs in response to differences in pressure. When the air pressure within the alveolar spaces falls below atmospheric pressure, air enters the lungs (inspiration), provided the larynx is open; when the air pressure within the alveoli exceeds atmospheric pressure, air is blown from the lungs (expiration). The flow of air is rapid or slow in proportion to the magnitude of the pressure difference. This explains the sciene of respiration and the mechanics of breathing.
Alveolar pressure fluctuations in the lungs resulting from tensing and relaxing of the muscles of the chest and abdomen. Each small increment of expansion transiently increases the space enclosing lung air. A difference in air pressure between atmosphere and lungs is created, and air flows in until equilibrium with atmospheric pressure is restored at a higher lung volume. When the muscles of inspiration relax, the volume of chest and lungs decreases, lung air becomes transiently compressed, its pressure rises above atmospheric pressure, and flow into the atmosphere results until pressure equilibrium is reached at the original lung volume. This, then, is the sequence of events during each normal respiratory cycle: lung volume change leading to the pressure difference, resulting in the flow of air into or out of the lung and establishment of a new lung volume.
The Lung–chest System
The forces that normally cause changes in the volume of the chest and lungs stem not only from muscle contraction but from the elastic properties of both the lung and the chest. A lung is like a balloon in that it resists stretch, tending to collapse almost totally unless held inflated by a pressure difference between its inside and outside. This tendency of the lung to collapse or pull away from the chest can be measured by carefully placing a blunt needle between the outside of the lung and the inside of the chest wall, thereby allowing the lung to separate from the chest at this particular spot. Force increases (pleural pressure becomes more negative) as the lung is stretched and its volume increases during inspiration. The force also increases in proportion to the rapidity with which air is drawn into the lung and decreases in proportion to the force with which air is expelled from the lungs. In summary, the pleural pressure reflects primarily two forces: (1) the force required to keep the lung inflated against its elastic recoil and (2) the force required to cause airflow in and out of the lung. Because the pleural pressure is below atmospheric pressure, the air is sucked into the chest and the lung collapses (pneumothorax) when the chest wall is perforated, as by a wound or by a surgical incision.
The force required to maintain inflation of the lung and to cause airflow is provided by the chest and diaphragm (the muscular partition between chest and abdomen), which are in turn stretched inward by the pull of the lungs. The lung–chest system thus acts as two opposed coiled springs, the length of each of which is affected by the other. Were it not for the outward traction of the chest on the lungs, these would collapse; and were it not for the inward traction of the lungs on the chest and diaphragm, the chest would expand to a larger size and the diaphragm would fall from its dome-shaped position within the chest.
The Role of Respiratory Muscles
The respiratory muscles displace the equilibrium of elastic forces in the lung and chest in one direction or the other by adding muscular contraction. During inspiration, muscle contraction is added to the outward elastic force of the chest to increase the traction on the lung required for its additional stretch. When these muscles relax, the additional retraction of the lung returns the system to its equilibrium position.
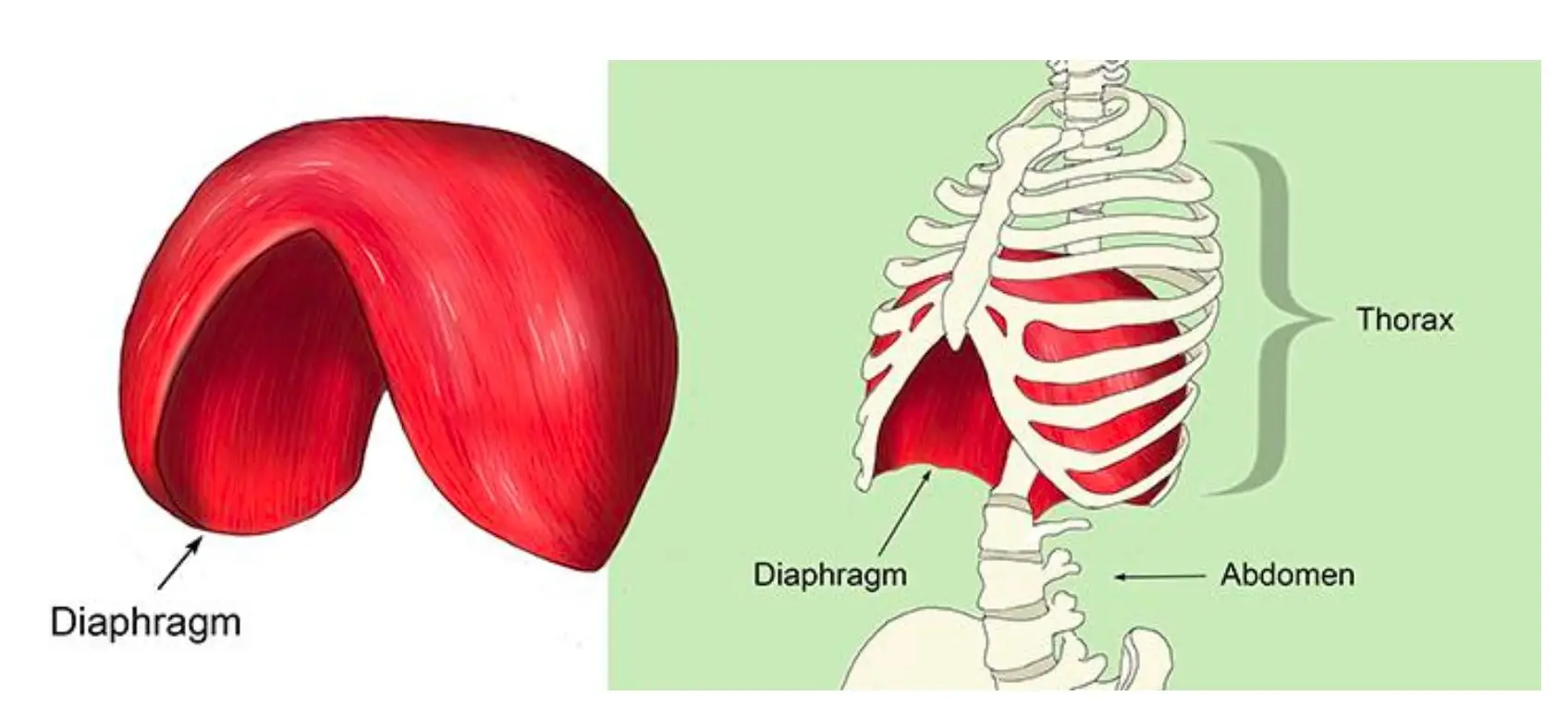
Contraction of the abdominal muscles displaces the equilibrium in the opposite direction by adding increased abdominal pressure to the retraction of lungs, thereby further raising the diaphragm, and causing forceful expiration. This additional muscular force is removed on relaxation and the original lung volume is restored. During ordinary breathing, muscular contraction occurs only on inspiration, expiration being accomplished “passively” by elastic recoil of the lung.
With total relaxation of the muscles of inspiration and expiration, the lung is distended to a volume—called the functional residual capacity—of about 40 percent of its maximum volume at the end of full inspiration. Further reduction of the lung volume results from the maximal contraction of the expiratory muscles of the chest and abdomen. The volume in these circumstances is known as the residual volume; it is about 20 percent of the volume at the end of full inspiration (known as the total lung capacity). The additional collapse of the lung to its “minimal air” can be accomplished only by opening the chest wall and creating a pneumothorax (or, collapsed lung).
The membranes of the surface of the lung (visceral pleura) and on the inside of the chest (parietal pleura) are normally kept close (despite the pull of lung and chest in opposite directions) by surface tension of the thin layer of fluid covering these surfaces. The strength of this bond can be appreciated by the attempt to pull apart two smooth surfaces, such as pieces of glass, separated by a film of water.
The Respiratory Pump and its Performance
The energy expended on breathing is used primarily in stretching the lung–chest system and thus causing airflow. It normally amounts to 1 percent of the baseline energy requirements of the body but rises substantially during exercise or illness. The respiratory pump is versatile, capable of increasing its output 25 times, from a normal resting level of about six liters (366 cubic inches) per minute to 150 liters per minute in adults. Pressures within the lungs can be raised to 130 centimeters of water (about 1.8 pounds per square inch) by the so-called Valsalva maneuver— i.e., a forceful contraction of the chest and abdominal muscles against a closed glottis (i.e., with no space between the vocal cords). Airflow velocity, normally reaching 30 liters per minute in quiet breathing, can be raised voluntarily to 400 liters per minute. Cough is accomplished by suddenly opening the larynx during a brief Valsalva maneuver. The resultant high-speed jet of air is an effective means of clearing the airways of excessive secretions or foreign particles. The beating of cilia (hairline projections) from cells lining the airways normally maintains a steady flow of secretions toward the nose, cough resulting only when this action cannot keep pace with the rate at which secretions are produced.
An infant takes 33 breaths per minute with a tidal volume (the amount of air breathed in and out in one cycle) of 15 milliliters, totaling about 0.5 liter— approximately one pint—per minute as compared to adult values of 14 breaths, 500 milliliters, and seven liters, respectively.
If the force of surface tension is responsible for the adherence of parietal and visceral pleurae, it is reasonable to question what keeps the lungs’ alveolar walls (also fluid-covered) from sticking together and thus eliminating alveolar airspaces. Such adherence occasionally does occur and is one of the complications of premature births. Normal lungs, however, contain a substance—a phospholipid surfactant—that reduces surface tension and keeps alveolar walls separated.
Gas Exchange
Respiratory gases—oxygen and carbon dioxide—move between the air and the blood across the respiratory exchange surfaces in the lungs. The structure of the human lung provides an immense internal surface that facilitates gas exchange between the alveoli and the blood in the pulmonary capillaries. The area of the alveolar surface in the adult human is about 50–100 square meters. Gas exchange across the membranous barrier between the alveoli and capillaries is enhanced by the thin nature of the membrane.
Respiratory gases move between the environment and the respiring tissues by two principal mechanisms, convection, and diffusion. Convection, or mass flow, is responsible for the movement of air from the environment into the lungs and for movement of blood between the lungs and the tissues. Respiratory gases also move by diffusion across tissue barriers such as membranes. Diffusion is the primary mode of transport of gases between air and blood in the lungs and between blood and respiring tissues in the body. The process of diffusion is driven by the difference in the partial pressures of gas between two locales. In a mixture of gases, the partial pressure of each gas is directly proportional to its concentration. The partial pressure of a gas in a fluid is a measure of its tendency to leave the fluid when exposed to a gas or fluid that does not contain that gas. A gas will diffuse from an area of greater partial pressure to an area of lower partial pressure regardless of the distribution of the partial pressures of other gases. There are large changes in the partial pressures of oxygen and carbon dioxide as these gases move between air and the respiring tissues. The partial pressure of carbon dioxide in this pathway is lower than the partial pressure of oxygen, due to different modes of transport in the blood, but almost equal quantities of the two gases are involved in metabolism and gas exchange.
Oxygen and carbon dioxide are transported between tissue cells and the lungs by the blood. The quantity transported is determined both by the rapidity with which the blood circulates and the concentrations of gases in the blood. The rapidity of circulation is determined by the output of the heart, which in turn is responsive to overall body requirements. Local flows can be increased selectively, as it occurs. One example is the flow through skeletal muscles during exercise. The performance of the heart and circulatory regulation is, therefore, important determinants of gas transport.
Oxygen and carbon dioxide are too poorly soluble in blood to be adequately transported in solution. Specialized systems for each gas have evolved to increase the quantities of those gases that can be transported in blood. These systems are present mainly in the red blood cells, which make up 40 to 50 percent of the blood volume in most mammals. Plasma, the cell-free liquid portion of blood, plays little role in oxygen exchange but is essential to carbon dioxide exchange.
Transport of Oxygen
Oxygen is poorly soluble in plasma so that less than 2 percent of oxygen is transported dissolved in plasma. Most of the oxygen is bound to hemoglobin, a protein contained within red cells. Hemoglobin is composed of four iron-containing ring structures (hemes) chemically bonded to a large protein (globin). Each iron atom can bind and then release an oxygen molecule. Enough hemoglobin is present in normal human blood to permit transport of about 0.2 mL of oxygen per mL of blood. The quantity of oxygen bound to hemoglobin is dependent on the partial pressure of oxygen in the lung to which blood is exposed. The curve representing the content of oxygen in blood at various partial pressures of oxygen, called the oxygen-dissociation curve, is a characteristic S-shape because binding of oxygen to one iron atom influences the ability of oxygen to bind to other iron sites. In alveoli at sea level, the partial pressure of oxygen is sufficient to bind oxygen to essentially all available iron sites on the hemoglobin molecule.
Hemoglobin is a protein made up of four polypeptide chains (α1, α2, β1, and β2). Each chain is attached to a heme group composed of porphyrin (an organic ring- like compound) attached to an iron atom. These iron-porphyrin complexes coordinate oxygen molecules reversibly, an ability directly related to the role of hemoglobin in oxygen transport in the blood.
Transport of Carbon Dioxide
The transport of carbon dioxide in the blood is considerably more complex. A small portion of carbon dioxide, about 5 percent, remains unchanged and is transported dissolved in the blood. The remainder is found in reversible chemical combinations in red blood cells or plasma. Some carbon dioxide binds to blood proteins, principally hemoglobin, to form a compound known as carbamate. About 88 percent of carbon dioxide in the blood is in the form of bicarbonate ion.
Less than 10 percent of the total quantity of carbon dioxide carried in the blood is eliminated during passage through the lungs. Complete elimination would lead to large changes in acidity between arterial and venous blood. Furthermore, blood normally remains in the pulmonary capillaries less than a second, an insufficient time to eliminate all carbon dioxide.
Only 5 percent of carbon dioxide in the blood is transported free in physical solution without chemical change or binding, yet this pool is important because only free carbon dioxide easily crosses biologic membranes. Virtually every molecule of carbon dioxide produced by metabolism must exist in the free form as it enters blood in the tissues and leaves capillaries in the lung. Between these two events, most carbon dioxide is transported as bicarbonate or carbamate.
Gas Exchange in the Lung
The introduction of air into the alveoli allows the removal of carbon dioxide and the addition of oxygen to venous blood. Because ventilation is a cyclic phenomenon that occurs through a system of conducting airways, not all inspired air participates in gas exchange. A portion of the inspired breath remains in the conducting airways and does not reach the alveoli where gas exchange occurs. This portion is approximately one-third of each breath at rest but decreases to as little as 10 percent during exercise, due to the increased size of inspired breaths.
In contrast to the cyclic nature of ventilation, blood flow through the lung is continuous, and almost all blood entering the lungs participates in gas exchange. The efficiency of gas exchange is critically dependent on the uniform distribution of blood flow and inspired air throughout the lungs. In health, ventilation and blood flow are extremely well matched in each exchange unit throughout the lungs. The lower parts of the lung receive slightly more blood flow than ventilation because gravity has a greater effect on the distribution of blood than on the distribution of inspired air. Under ideal circumstances, partial pressures of oxygen and carbon dioxide in alveolar gas and arterial blood are identical. Normally there is a small difference between oxygen tensions in alveolar gas and arterial blood because of the effect of gravity on matching and the addition of a small amount of venous drainage to the bloodstream after it has left the lungs. These events have no measurable effect on carbon dioxide partial pressures because the difference between arterial and venous blood is so small.
When Lungs don’t Function Optimally: Abnormal Gas Exchange
Lung disease can lead to severe abnormalities in blood gas composition. Because of the differences in oxygen and carbon dioxide transport, impaired oxygen exchange is far more common than impaired carbon dioxide exchange. Mechanisms of abnormal gas exchange are grouped into four categories— hypoventilation, shunting, ventilation–blood flow imbalance, and limitations of diffusion.
If the quantity of inspired air entering the lungs is less than is needed to maintain normal exchange—a condition known as hypoventilation—the alveolar partial pressure of carbon dioxide rises and the partial pressure of oxygen falls almost reciprocally. Similar changes occur in arterial blood partial pressures because the composition of alveolar gas determines gas partial pressures in blood perfusing the lungs. This abnormality leads to parallel changes in both gas and blood and is the only abnormality in gas exchange that does not cause an increase in the normally small difference between arterial and alveolar partial pressures of oxygen.
In shunting, venous blood enters the bloodstream without passing through functioning lung tissue. Shunting of blood may result from abnormal vascular (blood vessel) communications or blood flowing through unventilated portions of the lung (e.g., alveoli filled with fluid or inflammatory material). A reduction in arterial blood oxygenation is seen with shunting, but the level of carbon dioxide in arterial blood is not elevated even though the shunted blood contains more carbon dioxide than arterial blood.
The differing effects of shunting on oxygen and carbon dioxide partial pressures are the result of the different configurations of the blood-dissociation curves of the two gases. As noted before, the oxygen-dissociation curve is S-shaped and plateaus near the normal alveolar oxygen partial pressure, but the carbon dioxide-dissociation curve is steeper and does not plateau as the partial pressure of carbon dioxide increases. When blood perfusing the collapsed, unventilated area of the lung leaves the lung without exchanging oxygen or carbon dioxide, the content of carbon dioxide is greater than the normal carbon dioxide content. The remaining healthy portion of the lung receives both its usual ventilation and the ventilation that normally would be directed to the abnormal lung. This lowers the partial pressure of carbon dioxide in the alveoli of the normal area of the lung. As a result, blood leaving the healthy portion of the lung has a lower carbon dioxide content than normal. The lower carbon dioxide content in this blood counteracts the addition of blood with a higher carbon dioxide content from the abnormal area, and the composite arterial blood carbon dioxide content remains normal. This compensatory mechanism is less efficient than normal carbon dioxide exchange and requires a modest increase in overall ventilation, which is usually achieved without difficulty. Because the carbon dioxide- dissociation curve is steep and relatively linear, compensation for decreased carbon dioxide exchange in one portion of the lung can be counterbalanced by increased excretion of carbon dioxide in another area of the lung.
In contrast, the shunting of venous blood has a substantial effect on arterial blood oxygen content and partial pressure. Blood leaving an unventilated area of the lung has an oxygen content that is less than the normal content.
In the healthy area of the lung, the increase in ventilation above normal raises the partial pressure of oxygen in the alveolar gas and, therefore, in the arterial blood. The oxygen-dissociation curve, however, reaches a plateau at the normal alveolar partial pressure, and an increase in blood partial pressure results in a negligible increase in oxygen content. A mixture of blood from this healthy portion of the lung (with normal oxygen content) and blood from the abnormal area of the lung (with decreased oxygen content) produces a composite arterial oxygen content that is less than the normal level. Thus, an area of healthy lung cannot counterbalance the effect of an abnormal portion of the lung on blood oxygenation because the oxygen-dissociation curve reaches a plateau at a normal alveolar partial pressure of oxygen. This effect on blood oxygenation is seen not only in shunting but in any abnormality that results in a localized reduction in blood oxygen content.
Mismatching of ventilation and blood flow is by far the most common cause of a decrease in the partial pressure of oxygen in blood. There are minimal changes in blood carbon dioxide content unless the degree of mismatch is extremely severe. Inspired air and blood flow normally are distributed uniformly, and each alveolus receives approximately equal quantities of both. As the matching of inspired air and blood flow deviates from the normal ratio of 1 to 1, alveoli become either overventilated or under-ventilated in relation to their blood flow. In alveoli that are overventilated, the amount of carbon dioxide eliminated is increased, which counteracts the fact that there is less carbon dioxide eliminated in the alveoli that are relatively under-ventilated.
Abnormal gas exchange can also involve limitation of diffusion of gases across the thin membrane separating the alveoli from the pulmonary capillaries. A variety of processes can interfere with this orderly exchange; for oxygen, these include increased thickness of the alveolar-capillary membrane, loss of surface area available for the diffusion of oxygen, a reduction in the alveolar partial pressure of oxygen required for diffusion, and decreased time available for exchange due to increased velocity of flow. These factors are usually grouped under the broad description of “diffusion limitation,” and any can cause an incomplete transfer of oxygen with a resultant reduction in blood oxygen content. There is no diffusion limitation of the exchange of carbon dioxide because this gas is more soluble than oxygen in the alveolar-capillary membrane, which facilitates carbon dioxide exchange. The complex reactions involved in carbon dioxide transport proceed with sufficient rapidity to avoid being a significant limiting factor in exchange.
Working Together – Respiration, Circulation, and Metabolism
The interplay of respiration, circulation, and metabolism is the key to the functioning of the respiratory system. Cells set the demand for oxygen uptake and carbon dioxide discharge, that is, for gas exchange in the lungs. The circulation of the blood links the sites of oxygen utilization and uptake. The proper functioning of the respiratory system depends on both the ability of the system to make functional adjustments to varying needs and the design features of the sequence of structures involved, which set the limit for respiration.
The main purpose of respiration is to provide oxygen to the cells at a rate adequate to satisfy their metabolic needs. This involves the transport of oxygen from the lung to the tissues through the circulation of blood. In medieval times, the heart was regarded as a furnace where the “fire of life” kept the blood boiling. Modern cell biology has unveiled the truth behind the metaphor. Each cell maintains a set of furnaces, the mitochondria, where, through the oxidation of foodstuffs such as glucose, the energetic needs of the cells are supplied. The precise object of respiration, therefore, is the supply of oxygen to the mitochondria.
Cell metabolism depends on energy derived from high-energy phosphates such as adenosine triphosphate (ATP), whose third phosphate bond can release a quantum of energy to fuel many cell processes, such as the contraction of muscle fiber proteins or the synthesis of protein molecules. In the process, ATP is degraded to adenosine diphosphate (ADP), a molecule with only two phosphate bonds. To recharge the molecule by adding the third phosphate group requires energy derived from the breakdown of foodstuffs or substrates. Two pathways are available: (1) anaerobic glycolysis, or fermentation, which operates in the absence of oxygen; and (2) aerobic metabolism, which requires oxygen and involves the mitochondria. The anaerobic pathway leads to acid waste products and is wasteful of resources: The breakdown of one molecule of glucose generates only two molecules of ATP. In contrast, aerobic metabolism has a higher yield (36 molecules of ATP per molecule of glucose) and results in “clean wastes”—water and carbon dioxide, which are easily eliminated from the body and are recycled by plants in the process of photosynthesis. For any sustained high-level cell activity, the aerobic metabolic pathway is therefore preferable. Since oxidative phosphorylation occurs only in mitochondria, and since each cell must produce its ATP (it cannot be imported), the number of mitochondria in a cell reflects its capacity for aerobic metabolism or its need for oxygen.
The supply of oxygen to the mitochondria at an adequate rate is a critical function of the respiratory system because the cells maintain only a limited store of high- energy phosphates and oxygen, whereas they usually have a reasonable supply of substrates in stock. If oxygen supply is interrupted for a few minutes, many cells, or even the organism, will die.
Oxygen is collected from environmental air, transferred to blood in the lungs, and transported by blood flow to the periphery of the cells where it is discharged to reach the mitochondria by diffusion. The transfer of oxygen to the mitochondria involves several structures and different modes of transports. It begins with ventilation of the lung, which is achieved by convection or mass flow of air through an ingeniously branched system of airways. In the most peripheral airways, ventilation of alveoli is completed by diffusion of oxygen through the air to the alveolar surface. The transfer of oxygen from alveolar air into the capillary blood occurs by diffusion across the tissue barrier; it is driven by the oxygen partial pressure difference between alveolar air and capillary blood and depends on the thickness and the surface area of the barrier. Convective transport by the blood depends on the blood flow rate (cardiac output) and on the oxygen capacity of the blood, which is determined by its content of hemoglobin in red blood cells. The last step is the diffusive discharge of oxygen from the capillaries into the tissue and cells, which is driven by the oxygen partial pressure difference and depends on the quantity of capillary blood in the tissue. In this process, the blood plays a central role and affects all transport steps: oxygen uptake in the lung, transport by blood flow, and discharge to the cells. Blood also serves as a carrier for both respiratory gases: oxygen, which is bound to hemoglobin in the red blood cells, and carbon dioxide, which is carried by both plasma and red blood cells and which also serves as a buffer for acid-base balance in blood and tissues.
Metabolism, or, more accurately, the metabolic rate of the cells, sets the demand for oxygen. At rest, a human consumes about 250 ml (about 15 cubic inches) of oxygen each minute. With exercise, this rate can be increased more than 10-fold in a normal healthy individual, but a highly trained athlete may achieve a more than 20-fold increase. As more and more muscle cells become engaged in doing work, the demand for ATP and oxygen increases linearly with the work rate. This is accompanied by increased cardiac output, essentially due to a higher heart rate, and by increased ventilation of the lungs; as a consequence, the oxygen partial pressure difference across the air–blood barrier increases and oxygen transfer by diffusion are augmented. These dynamic adjustments to the muscles’ needs occur up to a limit that is twice as high in the athlete as in the untrained individual. This range of possible oxidative metabolism from rest to maximal exercise is called the aerobic scope. The upper limit to oxygen consumption is not conferred by the ability of muscles to do work, but rather by the limited ability of the respiratory system to provide or utilize oxygen at a higher rate. Muscle can do more work, but beyond the aerobic scope, they must revert to anaerobic metabolism, with the result that waste products, mainly lactic acid, accumulate and limit the duration of work.
The limit to oxidative metabolism is therefore set by some features of the respiratory system, from the lung to the mitochondria. Knowing precisely what sets the limit is important for understanding respiration as a key vital process, but it is not straightforward, because of the complexity of the system. Much has been learned from comparative physiology and morphology, based on observations that oxygen consumption rates differ significantly among species. For example, the athletic species in nature, such as dogs or horses, have an aerobic scope more than twofold greater than that of other animals of the same size; this is called adaptive variation. Then, oxygen consumption per unit body mass increases as animals become smaller, so that a mouse consumes six times as much oxygen per gram of body mass as a cow, a feature called allometric variation. Furthermore, the aerobic scope can be increased by training in an individual, but this induced variation achieves at best a 50 percent difference between the untrained and the trained state, well below interspecies differences.
Within the aerobic scope, the adjustments are due to functional variation. For example, cardiac output is augmented by increasing heart rate. Mounting evidence indicates that the limit to oxidative metabolism is related to structural design features of the system. The total amount of mitochondria in skeletal muscle is strictly proportional to maximal oxygen consumption, in all types of variation. In training, the mitochondria increase in proportion to the augmented aerobic scope. Mitochondria set the demand for oxygen, and they seem to be able to consume up to 5 ml (0.3 cubic inches) of oxygen per minute and a gram of mitochondria. If energy (ATP) needs to be produced at a higher rate, the muscle cells make more mitochondria. It is thus possible that oxygen consumption is limited at the periphery, at the last step of aerobic metabolism. But it is also possible that more central parts of the respiratory system may set the limit to oxygen transport, mainly the heart, whose capacity to pump blood reaches a limit, both in terms of rate and of the size of the ventricles, which determines the volume of blood that can be pumped with each stroke.
Digestion
Digestion is the term used to describe the sequence by which food is broken down and chemically converted so that it can be absorbed by the cells of an organism and used to maintain vital bodily functions. We will now summarize the actions of the digestive process.
To sustain themselves, all organisms must obtain nutrients from the environment. Some nutrients serve as raw materials for the synthesis of cellular material; others (e.g., many vitamins) act as regulators of chemical reactions; and still others, upon oxidation, yield energy. Not all nutrients, however, are in a form suitable for immediate use by an organism; some must undergo physical and chemical changes before they can serve as energy or cell substance.
Through the act of eating or ingestion, nutrients are taken from the environment. Many nutrient molecules are so large and complex that they must be split into smaller molecules before they can be used by the organism. This process of breaking down food into molecular particles of usable size and content is called digestion. Unusable components are expelled from the organism by a process called egestion, or excretion. Some plants, many microorganisms, and all animals perform these three functions—ingestion, digestion, and egestion (often grouped under the term alimentation)—but, as expected, the details differ considerably from group to group.
The problems associated with nutrient intake and processing differ greatly depending on whether the organism is autotrophic or heterotrophic. Autotrophic organisms can manufacture the large energy-rich organic compounds necessary for life from simple inorganic raw materials; consequently, they require only simple nutrients from the environment. By contrast, heterotrophic organisms cannot manufacture complex organic compounds from simple inorganic ones, and so they must obtain preformed organic molecules directly from the environment.
Green plants constitute by far most the Earth’s autotrophic organisms. During the process of photosynthesis, plants use light energy to synthesize organic materials from carbon dioxide and water. Both compounds can be absorbed easily across the membranes of cells—in a typical land plant, carbon dioxide is absorbed from the air by leaf cells, and water is absorbed from the soil by root cells—and used directly in photosynthesis; i.e., neither of them requires digestion. The only other nutrients needed by most green plants are minerals such as nitrogen, phosphorus, and potassium, which also can be absorbed directly and require no digestion.
Ingestion
As already explained, the nutrients obtained by most green plants are small inorganic molecules that can move with relative ease across cell membranes. Heterotrophic organisms such as bacteria and fungi, which require organic nutrients yet lack adaptations for ingesting bulk food, also rely on direct absorption of small nutrient molecules. Molecules of carbohydrates, proteins, or lipids, however, are too large and complex to move easily across cell membranes. Bacteria and fungi circumvent this by secreting digestive enzymes onto the food material; these enzymes catalyze the splitting of the large molecules into smaller units that are then absorbed into the cells. In other words, the bacteria and fungi perform extracellular digestion—digestion outside cells— before ingesting the food. This is often referred to as osmotrophic nutrition.
Like bacteria, protozoans are unicellular organisms, but their method of feeding is quite different. They ingest relatively large particles of food and carry out intracellular digestion (digestion inside cells) through a method of feeding called phagotrophic nutrition. Many protozoans also are osmotrophic to a lesser degree. Some organisms, such as amoebas, have pseudopodia (“false feet”) that flow around the food particle until it is completely enclosed in a membrane-bounded chamber called a food vacuole; this process is called phagocytosis. Other protozoans, such as paramecia, pinch off food vacuoles from the end of a prominent oral groove into which food particles are drawn by the beating of numerous small hair-like projections called cilia. In still other cases of phagotrophic nutrition, tiny particles of food adhere to the membranous surface of the cell, which then folds inward and is pinched off as a vacuole; this process is called pinocytosis. The food particles contained in vacuoles formed through phagocytosis or pinocytosis have not entered the cell in the fullest sense until they have been digested into molecules able to cross the membrane of the vacuole and become incorporated into the cellular substance. This is accomplished by enzyme- containing organelles called lysosomes, which fuse with the vacuoles and convert food into simpler compounds (see figure).
Most multicellular animals possess some sort of digestive cavity—a chamber opening to the exterior via a mouth—in which digestion takes place. The higher animals, including the vertebrates, have more elaborate digestive tracts, or alimentary canals, through which food passes. In all these systems, large particles of food are broken down into units of more manageable size within the cavity before being taken into cells and reassembled (or assimilated) as cellular substance.
Digestion
The enzymatic splitting of large and complex molecules into smaller ones is effective only if the enzyme molecules come into direct contact with the molecules of the material they are to digest. In animals that ingest very large pieces of food, only the molecules at the surface are exposed to the digestive enzymes. Digestion can proceed more efficiently, therefore, if the bulk food is first mechanically broken down, exposing more molecules for digestion. Among the variety of devices that have evolved to perform such mechanical processing of food are the teeth of mammals and the muscular gizzards of birds. Human digestion begins in the mouth. There, food is chewed and mixed with saliva, which adds moisture and contains the enzyme amylase, which begins to break down starches. The tongue kneads food into a smooth ball (bolus), which is then swallowed. The bolus passes through the pharynx and esophagus into the stomach, propelled by peristaltic muscular contractions. In the stomach, the food is then mixed by peristaltic contractions (about three per minute) with highly acidic gastric juices secreted there. The hormone gastrin stimulates the secretion of these juices, which contain water, inorganic salts, hydrochloric acid, mucin, and several enzymes. The food, now in a semiliquid state called chyme, passes from the stomach into the duodenum, the first section of the small intestine, where the greatest part of digestion takes place.
The chemical reactions involved in digestion can be clarified by an account of the digestion of maltose sugar. Maltose is, technically, a double sugar, since it is composed of two molecules of the simple sugar glucose bonded together. The digestive enzyme maltase catalyzes a reaction in which a molecule of water is inserted at the point at which the two glucose units are linked, thereby disconnecting them, as illustrated below.
In chemical terms, the maltose has been hydrolyzed. All digestive enzymes act similarly and thus are hydrolyzing enzymes.
Many other nutrient molecules are much more complex, being polymers, or long chains of simple component units. Starch, for example, is a carbohydrate, like maltose, but its molecules are composed of thousands of glucose units bonded together. Even so, the digestion of starch is essentially the same as the digestion of maltose: each linkage between adjacent glucose units is hydrolyzed, with the result that the starch molecule is split into thousands of glucose molecules. Protein molecules also are polymers, but their constituent units are amino acids instead of simple sugars. Proteolytic (i.e., protein-digesting) enzymes split the protein chains by hydrolyzing the bonds between adjacent amino acids. Because as many as 20 different kinds of amino acids may act as building blocks for proteins, the complete digestion of a protein into its amino acids requires the action of several different proteolytic enzymes, each capable of hydrolyzing the bonds between pairs of amino acids. Fat molecules too are composed of smaller building-block units (the alcohol glycerol plus three fatty acid groups); they are hydrolyzed by the enzyme lipase.
Various other classes of compounds are digested by hydrolytic enzymes specific to them. Not all these enzymes occur in every organism; for example, few animals possess cellulase (cellulose-digesting enzyme), even though cellulose constitutes much of the total bulk of the food ingested by plant-eating animals. Some nonetheless benefit from the cellulose in their diet because their digestive tracts contain microorganisms (known as symbionts) capable of digesting cellulose. The herbivores absorb some of the products of their symbionts’ digestive activity.
So far, the emphasis has been placed on the role of digestion in converting large complex molecules into smaller simpler ones that can move across membranes, which thus permits the absorption of food into cells. The same processes occur when substances must be moved from cell to cell within a multicellular organism. Thus, green plants, which do not have to digest incoming nutrients, digest stored material, such as starch, before it can be transported from storage organs (tubers, bulbs, corms) to points of utilization, such as growing buds.
The Liver
We could probably write an entire chapter dedicated entirely to the liver and its functions.
The liver is responsible for detoxifying various metabolites; it also synthesizes proteins and produces biochemicals necessary for digestion and growth. Located in the right upper quadrant of the abdomen, the liver holds about one pint (13%) of the body’s blood supply at any given moment. The liver consists of 2 main lobes. Both are made up of 8 segments that consist of nearly 1,000 smaller lobes (lobules). These lobules are connected to small ducts (tubes) that connect with larger ducts to form the common hepatic duct. The common hepatic duct transports the bile made by the liver cells to the gallbladder and duodenum (the first part of the small intestine) via the common bile duct.
The liver regulates most chemical levels in the blood and excretes a product called bile. This helps carry away waste products from the liver. All the blood leaving the stomach and intestines passes through the liver. The liver processes this blood and breaks down, balances, and creates the nutrients and also metabolizes drugs into forms that are easier to use for the rest of the body or that are nontoxic. More than 500 vital functions have been identified with the liver. Some of the more well- known functions are included in the discussion that follows.
The production of bile is important, as it aids in carrying away waste and break down fats in the small intestine during digestion. The liver is responsible for the production of certain proteins (for blood plasma), cholesterol and special proteins to help carry fats through the body. The liver converts excess glucose into glycogen for storage (glycogen can later be converted back to glucose for energy) and to balance and make glucose as needed.
The liver regulates blood levels of amino acids, the building blocks of proteins, and processes hemoglobin for use of its iron content (the liver stores iron). The conversion of poisonous ammonia to urea (urea is an end product of protein metabolism and is excreted in the urine) is done by the liver as well.
The liver also helps the body detoxify itself, ridding the blood of drugs and other poisonous substances. In addition to regulating blood clotting, the liver helps the body to resist infections by making immune factors and removing bacteria from the bloodstream.
When the liver has broken down harmful substances, its by-products are excreted into the bile or blood. Blood by-products are filtered out by the kidneys and leave the body in the form of urine.
The Kidneys
The kidneys are a pair of kidney bean-shaped organs located just above the waist, between the peritoneum and the back of the abdomen. The two kidneys lie behind the liver and the intestines in the small of the back. They are partially protected by the 11th and 12th pairs of ribs. The left kidney is slightly higher than the right, since the liver occupies the space above the kidney on the right side.
The kidneys are the hardest working organs of the urinary system. The other components of the urinary system serve mainly as passageways or for the storage of urine. The functions of the kidneys include: excreting wastes and foreign substances in the urine while regulating various properties of blood, including:

Functions of the kidneys include the reabsorption of solutes and water, production of urine, and excretion of wastes. The kidneys also perform several homeostatic functions, including regulation of electrolytes, acid-base balance, and blood pressure. They also produce the hormones vitamin D.
The Nervous System
In humans, the nervous systems consist of two main parts, called the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS contains the brain and spinal cord. The PNS consists mainly of nerves, which are long fibers that connect the CNS to every other part of the body but also includes other components such as peripheral ganglia, sympathetic and parasympathetic ganglia, and the enteric nervous system, a semi- independent part of the nervous system whose function is to control the gastrointestinal system.
At the cellular level, the nervous system is defined by the presence of a special type of cell, called the neuron, also known as a “nerve cell”. Neurons have special properties that allow them to send signals rapidly and precisely to other cells. They send these signals in the form of electrochemical waves traveling along thin fibers called axons, which causes chemicals called neurotransmitters to be released at junctions to other neurons, called synapses. A cell that receives a synaptic signal from a neuron (a postsynaptic neuron) may be excited, inhibited, or otherwise modulated. The connections between neurons form neural circuits that can generate very complex patterns of dynamical activity. Along with neurons, the nervous system also contains other specialized cells called glial cells (or simply glia), which provide structural and metabolic support. Recent evidence suggests that glia may also have a substantial signaling role.
At the most basic level, the function of the nervous system is to control the movement of the organism and to affect the environment (e.g., through pheromones). This is achieved by sending signals from one cell to others, or from one part of the body to others. The output from the nervous system derives from signals that travel to muscle cells, causing muscles to be activated, and from signals that travel to endocrine cells, causing hormones to be released into the bloodstream or other internal fluids. The input to the nervous system derives from sensory cells of widely varying types, which transmute physical modalities such as light and sound into neural activity. Internally, the nervous system contains complex webs of connections between nerve cells that allow it to generate patterns of activity that depend only partly on sensory input. The nervous system is also capable of storing information over time, by dynamically modifying the strength of connections between neurons, as well as other mechanisms The Endocrine System
The endocrine system includes the areas in the body that regulate and produce hormones. Hormones are the messengers that allow our organs and cells to communicate to produce a coordinated effect. Without our endocrine system, various parts of the body would work independently from one another. Some of the organs that produce hormones include the pineal gland (which sets the circadian rhythm), the thyroid (which sets cellular metabolism), the pancreas (which is involved in digestion and blood sugar control), the ovaries and testes (which produce the sex hormones estrogen, progesterone, and testosterone), and the adrenal glands (which produce cortisol that balances emotional and physiological stress to maintain homeostasis). Hormone levels fluctuate throughout the day and are released in a pulse-like manner throughout the hour, day, or month. This release is set by the circadian rhythm. Melatonin can be considered the re-set button for the circadian rhythm and it allows the body to recover and repair from work done during the day. Melatonin is produced during the stage of deep sleep, making good healthy sleep critical for balancing the endocrine system.
What are the top factors that throw off balance within the endocrine system?
- Poor quality or inadequate sleep.
- Environmental pollutants such as exhaust fumes, pesticides, paint fumes, heavy metals, plastics, unfiltered drinking water, smoking, and estrogens found in water bottles and some makeup.
- A lack of Vitamin D from staying in the office too long and not getting out in the sun. Vitamin D is a potent modulator of the endocrine system, especially with the sex hormones.
- Emotions – the production and release of our neurotransmitters (GABA, serotonin, dopamine, etc.) are linked with the endocrine hormones. An imbalance in one system, whether it’s insufficient production of a neurotransmitter like serotonin, can throw off the balance of the other system.
- Stress – whether emotional or physical such as from chronic pain like endometriosis will cause a consistent rise in cortisol from the adrenals. This will alter neurotransmitter production (affecting your emotions), it will deplete nutrients needed to produce other hormones and neurotransmitters, it will steal the production line away from other hormones and shunt the production supplies to making cortisol (creating imbalances in hormones such as estrogen and progesterone), and it burns through key nutrients such as the B vitamins, zinc, and Vitamin C. A stress is more than just emotional stress, it can also be from physical stress such as from persistent low blood sugar, chronic pain, chronic disease, inflammation, food allergies, and imbalances in the gut.
Holistically speaking, the endocrine system is vast, including every hormone and its functions within the body. Therefore, we will place more of an emphasis on those that are more prominent to be aware of for the CHNC. This group will include discussion on the following;
- growth hormone
- thyroid hormones
- insulin
- corticosteroids
Growth Hormone
Also known as somatropin, human growth hormone (HGH) Increases gene expression, protein synthesis, fat breakdown, and bone synthesis. hGH is a natural hormone and is produced in the human body by the pituitary gland. Like all other hormones, it is a messenger. If levels are too low or too high, there are undesirable effects. A human growth hormone deficiency can cause damage to the body and its systems. This is how hormone affect hunger.
In recent years, human growth hormone abuse has had heavy media coverage. We are not placing an emphasis on the supplementation of hGH; there is too much debate within the scientific community on the matter. The media and popular culture have contributed greatly to the confusion regarding this hormone and these are diagnosed only by a physician; this is one example of an area the CHNC should avoid as it is mired in controversy and is often the subject of misinformation. We need to know its relevance, but we aren’t drawing blood and sending it to a lab for analysis. We leave hGH therapies for health and medical professionals. But be prepared to have a dialogue with clients who have questions – and be prepared to answer questions only for which you have facts. It is okay to inform your client that this is best left to a medical professional. Crossing the line with recommendations has led to many problems in the coaching industry. We can avoid this, and we should.
Thyroid Hormones
- At normal levels—critical to maintaining healthy bone metabolism
- At elevated levels, (hyperthyroidism) there is a marked increase in bone resorption, leading to bone loss
- Levels below normal (hypothyroidism) result in a slowing of bone turnover and diminishes the bone calcium pool
Of the 20 million Americans who have thyroid disease, most have hypothyroidism, a condition where the thyroid gland can’t make enough thyroid hormone to keep the body running normally.
Low thyroid levels are more common in women, hypothyroidism can have many genetic, nutritional, and immune-related underlying causes and contributing factors.
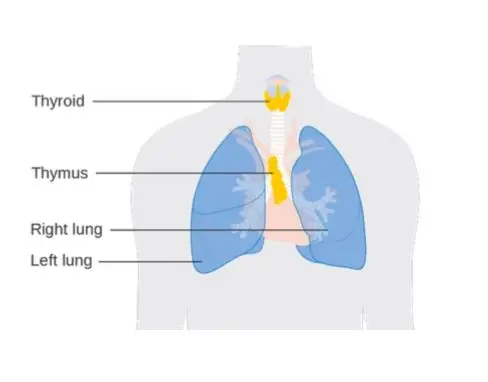
A Naturopath may address hypothyroidism from the root of the problem. There are two main types of hypothyroidism, with numerous contributing factors:
Autoimmune thyroid disease (Hashimoto’s) is the most common cause of hypothyroidism. This is a form of thyroid inflammation caused by the patient’s immune system. Hashimoto’s is five to eight times more common in women than men. Elevated levels of anti-TPO antibodies are found in approximately five percent of adults and 15 percent of older women. Individuals with other autoimmune diseases such as type 1 diabetes, celiac disease, and rheumatoid arthritis have a higher prevalence of autoimmune thyroid disease.
With Central or pituitary hypothyroidism, the thyroid just isn’t making enough hormone, is often influenced by environmental and nutritional factors including gastrointestinal system inflammation, vitamin and mineral deficiencies or imbalances, high levels of cortisol (due to prolonged stress or oral steroid use), and/or elevated estrogen. Surgery on the thyroid gland (to remove a goiter or nodule) and/or radioactive iodine treatment (to treat a thyroid nodule, hyperactive thyroid, throat cancer, and cancer of nearby sites) can also cause hypothyroidism.
We must provide caution when describing Naturopathic Doctors. There is a considerable difference between one who identifies as a “Naturopath’ as compared to an ND (Doctor of Naturopathic). You will want to be familiar with these distinctions, more can be found in your Research Articles section within this course.
- Vitamin B-12 is very important for thyroid production. It helps improve cellular response to thyroid hormone and boosts energy production in cells to help with fatigue and other symptoms associated with hypothyroidism.
- Selenium supports efficient thyroid synthesis and metabolism and has been found in studies to reduce thyroid antibody levels in patients with Hashimoto’s.
- Zinc is beneficial in improving thyroid function and hormone levels and has been shown in studies to have a positive effect on thyroid function in overweight females.
Botanical Herbs
Several herbs have been shown to benefit thyroid function, including adaptogens like Ashwagandha, which was shown to improve thyroid function for subclinical hypothyroid patients. Adaptogens are herbs that work on several body functions at once to help balance out dysfunction with regulating metabolic processes. They are used with caution, however, in those with autoimmune thyroid disease as some botanicals can stimulate the immune system response. Other herbs such as bum guggal have been used in all forms of hypothyroidism as they are believed to help to convert the inactive form of T4 to the more active form of T3. Any herbal treatment should be approved by a professional trained in herbal medicine to reduce side effects, prevent unwanted drug/herb interactions, and maximize treatment efficacy.
Insulin
Obesity is a pandemic in our world, especially abdominal obesity. While obesity is highly correlated with abnormal biochemical profiles (high blood lipids, insulin, and glucose), lower body obesity is often considered relatively less dangerous in terms of health. It is not, however, truly benign. However, lower body obesity can also contribute to increased non-esterified free fatty acids (NEFAs). Obesity in the lower body can also stress the weight-bearing joints, contribute to varicose veins and osteoarthritis, and discourage participation in regular physical activity due to the discomfort associated with carrying excess body weight. You may need to modify training for clients with diabetes and insulin sensitivity.
Once BMI reaches a certain critical point, however, mortality increases. Mortality increases as BMI increases above 25, with the sharpest risk associated with a BMI >30. From this curve, one would also conclude that the risk of mortality increases at a BMI <20 – as this would be underweight.
Corticosteroids
Corticosteroids are a class of drug that lowers inflammation in the body. They also reduce immune system activity. Because corticosteroids ease swelling, itching, redness, and allergic reactions, doctors often prescribe them to help treat diseases like asthma and arthritis. Our bodies are equipped with an intelligent immune system that works to protect us against foreign invaders including bacteria, viruses and parasites. However, sometimes the immune system can malfunction and, instead of fighting against pathogens, it produces auto-antibodies that begin to attack the body itself. That said, corticosteroids are commonly used in modern medicine. But is there a natural approach available? Yes. Are they just as effective? Maybe not.
Some plant chemicals, also known as phytochemicals, act as potent anti- inflammatory agents by keeping inflammatory mediators – chemicals that are released from cells because of an inflammatory response – at bay. Spices such as turmeric and ginger root are rich in anti-inflammatory phytochemicals, and for this reason are great examples of easily accessible, natural anti-inflammatories.
Since curcumin, the main anti-inflammatory found in turmeric, is alleged to be more easily absorbed when combined with either black pepper or plant oils, try combining all three to create a delicious seasoning for curried chicken. Alternatively, a turmeric supplement can deliver a high-quality dose of fast-acting liquid curcumin in an easy-to-ingest capsule. However, phytochemicals such as pygnogenol and grape seed extract have spotty research findings.
Natural Corticosteroid Support: Licorice Root
Glycyrrhiza glabra, more commonly known as licorice root, contains a plant compound called glycyrrhizic acid, which elicits an anti-inflammatory effect by mimicking the actions of steroids in the body. It does this by inhibiting an enzyme that causes excessive activation of steroid receptors. Therefore licorice root may also be used during the process of weaning off a steroid medication. However, be aware that licorice can cause high blood pressure, so should only be taken under medical advice. Licorice root is often consumed in tea; the boiled root is steeped with licorice leaves to make an herbal tea.
